





Abstract
In December 2019, SARS-CoV-2 emerged causing the COVID-19 pandemic. SARS-CoV, the agent responsible for the 2003 SARS outbreak, utilises ACE2 and TMPRSS2 host molecules for viral entry. ACE2 and TMPRSS2 have recently been implicated in SARS-CoV-2 viral infection. Additional host molecules including ADAM17, cathepsin L, CD147, and GRP78 may also function as receptors for SARS-CoV-2.
To determine the expression and in situ localisation of candidate SARS-CoV-2 receptors in the respiratory mucosa, we analysed gene expression datasets from airway epithelial cells of 515 healthy subjects, gene promoter activity analysis using the FANTOM5 dataset containing 120 distinct sample types, single cell RNA sequencing (scRNAseq) of 10 healthy subjects, proteomic datasets, immunoblots on multiple airway epithelial cell types, and immunohistochemistry on 98 human lung samples.
We demonstrate absent to low ACE2 promoter activity in a variety of lung epithelial cell samples and low ACE2 gene expression in both microarray and scRNAseq datasets of epithelial cell populations. Consistent with gene expression, rare ACE2 protein expression was observed in the airway epithelium and alveoli of human lung, confirmed with proteomics. We present confirmatory evidence for the presence of TMPRSS2, CD147, and GRP78 protein in vitro in airway epithelial cells and confirm broad in situ protein expression of CD147 and GRP78 in the respiratory mucosa.
Collectively, our data suggest the presence of a mechanism dynamically regulating ACE2 expression in human lung, perhaps in periods of SARS-CoV-2 infection, and also suggest that alternate receptors for SARS-CoV-2 exist to facilitate initial host cell infection.
Abstract
ACE2 gene and protein expression is low to absent in airway and alveolar epithelial cells in human lungs. Our data suggest the presence of a mechanism dynamically regulating ACE2 expression in human lung or other receptors for SARS-CoV-2.
INTRODUCTION
In 2003, the severe acute respiratory syndrome (SARS) outbreak caused by the SARS coronavirus (CoV) resulted in 8096 probable cases with 774 confirmed deaths [1, 2] In patients with SARS, deaths were attributed to acute respiratory distress associated with diffuse bilateral pneumonia and alveolar damage [3]. In December 2019, SARS-CoV-2 emerged causing the COVID-19 pandemic. SARS-CoV-2 is spreading at a much more rapid rate than SARS-CoV [4–6]. Similar clinical reports of diffuse bilateral pneumonia and alveolar damage have been reported [7–9]. Severe cases of SARS-CoV-2 have been associated with infections of the lower respiratory tract with detection of the virus throughout this tissue as well as the upper respiratory tract [7–9]. The biological mechanisms that may govern differences in the number of SARS and COVID-19 cases remain undefined. It is possible that SARS-CoV-2 possesses distinct molecular mechanisms that impact the virulence through viral proteins, greater susceptibility of host cells to infection, permissivity of host cells to virus replication, or some combination of these and other potentially unknown factors [10–13]. Understanding SARS and SARS-CoV-2 virus similarities and differences at the molecular level in the host may provide insights into transmission, pathogenesis, and interventions.
The seminal report identifying the receptor for SARS-CoV used a HEK293 cell over-expression system to identify angiotensin-converting enzyme 2 (ACE2) as a receptor by co-immunoprecipitation with SARS-CoV spike domain 1 [14]. Subsequently, spike protein of SARS-CoV was identified as the viral interacting partner of ACE2. Host protease activity by TMPRSS2 facilitates ACE2 ectodomain cleavage and fusion of SARS-CoV membrane with host cell membrane [15–17]. ADAM17 has also been demonstrated to cleave ACE2 ectodomain, but this was not required for SARS CoV infection [18–20]. Mechanisms of SARS CoV entry distinct from ACE2 have also been reported and include activation by endosomal cathepsin L and cell surface expression of CD147 or GRP78 [21–23] Each of these receptors were mechanistically interrogated and suggest that SARS CoV could initiate host cell entry and infection using multiple mechanisms. Recent in vitro reports have demonstrated that similar host proteins are involved in facilitating cell entry by SARS-CoV-2, such as ACE2 and TMPRSS2 [5, 24] Biophysical and structural evidence strongly support an interaction of ACE2 with SARS-CoV-2 spike protein, similar to SARS-CoV spike protein [12, 13]. Molecular docking studies have also suggested that SARS-CoV-2 spike protein can interact with cell-surface GRP78 [25]. Indirect evidence for a role of CD147 in SARS-CoV-2 binding has been demonstrated in vitro with the use of an anti-CD147 intervention that prevented virus replication [26]. Furthermore, a clinical study with an anti-CD147 intervention reduced symptoms and duration of hospital admission for COVID-19 patients [27]. In summary, although there is evidence that SARS-CoV-2 and SARS-CoV both utilise ACE2 as a receptor to facilitate virus entry, it is possible that differences in host-entry mechanisms play a role in the large epidemiological differences between the two viruses, which may include additional unidentified receptors.
ACE2 and TMPRSS2 were identified as cellular entry determinants for SARS-CoV using mechanistic studies. The original report of in situ human lung ACE2 expression described positive immunohistochemical staining for alveoli and airway epithelial cells, and immunocytochemical staining in A549 type II alveolar epithelial cells [28]. ACE2 protein expression is also present in the human lung adenocarcinoma cell line, Calu-3 [29]. Similar to ACE2, the original report describing the expression of TMPRSS2 in human respiratory mucosa described expression in airway epithelium and type II alveolar epithelial cells [30]. The specificity of the ACE2 and TMPRSS2 antibodies used for analysis of expression patterns in human lung tissues remains to be addressed.
To address the uncertainties related to SARS-CoV-2 receptors in human lung, we performed gene expression and in situ protein profiling of candidate receptors in human airway epithelial cells and lung tissue. Our computational analysis used publicly available microarray gene expression datasets from airway epithelial cells of 515 unique subjects, single cell sequencing data from 10 subjects, and the FANTOM5 dataset for promoter activities of 74 lung-related cell and tissue types. Our proteomic analysis used data from the Human Proteome Map [31] and a dataset from primary human airway epithelial cells grown under air-liquid interface culture conditions [32]. For our in situ protein profiling, we performed immunohistochemical analysis of 98 human lung tissue samples. To determine antibody specificity, we performed immunoblots on protein isolated from Calu-3 cells, primary human airway epithelial cells, primary type II alveolar epithelial cells, the human bronchial epithelium cell (HBEC)-6KT cell line, the A549 type II alveolar epithelial cell line, and HEK cells. Collectively our data contrast previous reports, demonstrating rare ACE2 protein expression in the airway epithelium and alveoli of human lung. Our protein expression data are consistent with low ACE2 promoter activity in a variety of lung epithelial cell samples and low ACE2 gene expression in both microarray and single cell RNA sequencing (scRNAseq) datasets. We present confirmatory evidence for the presence of TMPRSS2, CD147, and GRP78 protein in vitro in airway epithelial cells and confirm broad in situ protein expression of CD147 and GRP78 in the respiratory mucosa. Our data suggest that the presence of a mechanism dynamically regulating ACE2 expression in human lung, perhaps in periods of SARS-CoV-2 infection and/or that alternate receptors for SARS-CoV-2 exist to facilitate initial host cell infection in lung tissue.
METHODS
Human ethics
Procurement of primary human airway epithelial cells used for immunoblots and lung tissue for immunohistochemistry was approved by Hamilton integrated Research Ethics Board (HiREB 5099T, 5305T, 11-3559 and 13-523-C). UBC Research Ethics Office approved heart tissue archives and primary human airway epithelial cell collection.
Upper and lower airway gene expression analysis
Public microarray experiments using Affymetrix chips (HuGene-1.0-st-v1 and HG-U133 Plus 2) on airway epithelial cell samples collected from nasal (GSE19190) or bronchial (GSE11906) brushings of healthy, non-smokers were obtained from the NCBI Gene Expression Omnibus (GEO) database [33, 34]. This resulted in a total of 80 individual samples from the two different experiments which includes 11 upper airway samples (Nasal: 11) and 69 lower airway samples (Trachea: 17, Large Airway: 17, Small Airway: 35). For all dataset samples, raw intensity values and annotation data were downloaded using the GEOquery R package (version 2.52.0) [35] from the Bioconductor project [36]. Probe definition files were downloaded from Bioconductor and probes were annotated using Bioconductor's annotate package. All gene expression data were unified into a single dataset that was then RMA-normalised, and only genes present in both of the Affymetrix platforms (N=16 013) were kept for subsequent analyses. Correction of experiment-specific batch effects was performed using the ComBat method [37] implemented using the sva R package (version 3.32.1) [38]. RMA-normalised expression levels for conventional (ACE2, TMPRSS2, ADAM17, and CTSL) and non-conventional (CD147, and GRP78) SARS-CoV-2 receptor genes were compared across the four defined airway levels, with CDH1 expression level included as a positive control with known expression in lung tissue. Gene expression levels were tested for significant differences via pairwise Wilcoxon rank sum tests with Benjamini-Hochberg multiple testing correction using the stats R package (version 3.6.1). Gene expression box plots were generated with the ggplot2 R package (version 3.2.1).
Analysis of Curated Bronchial Epithelial Cell Brushing Dataset
A total of 1859 public microarray experiments using Affymetrix chips (HG-U133 Plus 2 and HuGene-1.0-st-v1) on airway epithelial cell samples were selected from the NCBI GEO database. These samples were further filtered by removing individuals with asthma or COPD, resulting in a total of 504 individual healthy samples (GSE4302, 28 samples; GSE67472, 43 samples; GSE37147, 159 samples; GSE108134, 274 samples). Within this dataset, sex and age information was included for 310 samples with 86 females/106 males. Smoking status information was also provided for 451 samples with 260 current smokers, 82 former smokers, and 109 never smokers.
For all dataset samples, raw intensity values and annotation data were downloaded as described above. Probe definition files were retrieved as described above. All gene expression data were unified into a single dataset that was then RMA-normalised, and only genes present in both of the Affymetrix platforms (N=16 105) were kept for subsequent analyses. Correction of experiment-specific batch effects was performed as described above.
Analysis of promoter activity from the FANTOM5 dataset
The FANTOM5 promoterome dataset [39] for the hg38 assembly [40] was used to examine promoter activity of SARS-CoV-2-related human genes, namely ACE2, TMPRSS2, ADAM17, CTSL (cathepsin L1), CD147 and GRP78. Using the ZENBU genome browser [41], the nearest cap analysis of gene expression (CAGE) peak upstream and on the same strand as each of the aforementioned genes was extracted and analysed. The dataset consists of CAGE promoter activity data for 1886 primary cells, cell lines, and tissues from humans, and quantified as normalised transcripts per million (TPM). A subset of FANTOM5 CAGE data (120 samples) is presented considering only samples related to lung, gut, heart, and prostate tissues (consisting of 74, 19, 15, and 12 samples, respectively). Normalised TPM values for each CAGE peak, an approximation for promoter activity, were log10 transformed and separated according to tissue and cell type, and the radius of each point is proportional to these transformed normalised TPM values.
Analysis of protein abundance from proteomic datasets
Publicly available human proteomic data from Kim et al. [31] and Foster et al. [32] datasets were used to evaluate SARS-CoV-2 receptor-related protein expression in different human tissues and experimental conditions. Expression values were extracted from the Kim et al. dataset for ACE2, TMPRSS2, ADAM17, CTSL (cathepsin L1), BSG (CD147), HSPA5 (GRP78), using CDH1 as a control for airway cells. Data were created using the pheatmap package in R (version 1.0.12) and expressed as log10-transformed to facilitate visualisation. Proteomic data from the Foster et al. dataset consists of bronchial epithelial cells collected from healthy, non-smokers (N=4; male) and exposed to phosphate-buffered saline control vehicle. Intensity values for ACE2, TMPRSS2, ADAM17, CD147, and GRP78 were extracted with CDH1 included as a positive control. Intensity values were determined by the original study authors via normalisation of all detected peptide intensities associated with a given parent protein [32]. Box plots were generated with the ggplot2 R package (version 3.2.1) with intensity values log10-transformed for visualisation purposes.
Analysis of single cell RNA sequencing (scRNAseq) data
Data preprocessed using the Cell Ranger pipeline (10x Genomics) were obtained from GSE135893. Samples from 10 control subjects and 12 IPF patients were downloaded and post-processed with Seurat package in R [42]. Cell populations were defined using the markers provided in the source paper [43]. Cells belonging to the 10 control subjects were used for further analysis. Visualisations of violin plots were created using Seurat.
Primary Human Airway Epithelial Cells
The human lung adenocarcinoma cell line, Calu-3, was grown under culture conditions defined by the supplier (ATCC-HTB-55). Primary human airway epithelial cells isolated via bronchial brushings from consented healthy individuals were grown in PneumaCult ExPlus (Stemcell Technologies, Vancouver Canada) under submerged monolayer culture conditions and used in between passage 1 and 4. The human bronchial epithelial cell line, HBEC-6KT, was grown under submerged monolayer culture conditions in keratinocyte serum free media supplemented with epidermal growth factor (0.4 ng·mL−1) and bovine pituitary extract (50 μg·mL−1) [44–47].
Immunoblots
Cell protein was isolated using RIPA lysis buffer (VWR, Ontario, Canada) supplemented with protease inhibitor cocktail (Sigma, Ontario, Canada) with quantification performed using Bradford assay reagents (Bio Rad, Ontario, Canada). Immunoblots were performed using stain free 4–20% pre-cast gradient gels and imaged on a ChemiDoc XRS+ Imaging system (Bio Rad, Ontario, Canada). For each immunoblot, 20 μg of protein was added per lane. ACE2 (R&D Systems - MAB933 – Monoclonal - Clone 171 606 – 2 μg·mL−1), TMPRSS2 (Atlas Antibodies - HPA035787 – Polyclonal – 0.4 μg·mL−1), CD147 (Abcam – ab666 – Monoclonal – Clone MEM-M6/1 – 1 μg·mL−1), and GRP78 (BD – 610 979 – Monoclonal – Clone – 40/BiP – 0.25 μg·mL−1 and Atlas Antibodies – HPA038845 – Rabbit Polyclonal) primary antibodies were diluted in 5% skim milk/Tris buffered saline with 0.1% Tween-20 and incubated overnight on a rocker at 4°C with detection performed the following day using an anti-mouse-HRP (ACE2, CD147, GRP78 - BD) or anti-rabbit-HRP (TMPRSS2, GRP78 - Atlas) conjugated secondary antibodies at 1:3000 for 2 hrs at room temperature (Cell Signalling, Danvers, MA, USA). Visualisation of TMPRSS2, CD147, and GRP78 was performed using was performed using Clarity™ Western enhanced chemiluminescence (ECL) Substrate, while ACE2 was visualised with Clarity Max™ ECL Substrate (Bio Rad, Ontario Canada). Total protein loading images were collected as a qualitative visualisation of protein loading between sample types [48]. The immunogen for ACE2 primary antibody is mouse myeloma cell line NS0-derived recombinant human ACE2 Gln18-Ser740 (predicted). The immunogen for TMPRSS2 primary antibody is the recombinant protein epitope signature tag antigen sequence, GSPPAIGPYYENHGYQPENPYPAQPTVVPTVYEVHPAQYYPSPVPQYAPRVLTQASNPVVCTQPKSPSGTVCTSKT. The immunogen for the CD147 primary antibody is recombinant full-length protein corresponding to human CD147. The immunogen for the GRP78 - BD primary antibody is human BiP/GRP78 amino acids 525–628. The immunogen for the GRP78 - Atlas primary antibody is the recombinant protein epitope signature tag antigen sequence, EKFAEEDKKLKERIDTRNELESYAYSLKNQIGDKEKLGGKLSSEDKETME
KAVEEKIEWLESHQDADIEDFKAKKKELEEIVQPIISKL.
Independent immunoblot analysis (L.O, C.J. A.J. and G.J) were performed on A549, HEK, and immortalised human bronchial epithelial cells. Equal amounts of protein (20 µg) were loaded on to 4–12%, Bis-Tris gradient gels (ThermoFisher, NP0326BOX) with anti-ACE2 (Abcam – ab108252 – Rabbit monoclonal – Clone EPR4435(2) – 1/500 dilution of stock antibody). A loading control of GAPDH was used to demonstrate protein loading (Abcam – ab181603 – Rabbit monoclonal – EPR16884 – 1/10 000 dilution of stock antibody). Visualisation was performed with ECL Clarity (Biorad, UK) on a Licor C-DiGit.
Immunohistochemistry
Formalin-fixed paraffin-embedded human lung tissue from non-diseased regions were obtained from archived tissue blocks from patients who had undergone lung resection for clinical care. Human heart tissue was from the UBC Cardiovascular Tissue Registry. Four-micron thick sections were cut and stained for ACE2 (15 μg·mL−1), TMPRSS2 (10 μg·mL−1), CD147 (5 μg·mL−1), and GRP78 (HPA038845 – 1/200 dilution) using the same antibodies used for immunoblot analysis. All staining was performed on a Leica Bond RX system with Leica Bond reagents, heat-induced antigen retrieval in at pH 6 (20 min) with primary antibody incubation for 20 min. Digital slide scanning was performed using an Olympus VS120-L100 Virtual Slide System at 40X magnification with VS-ASW-L100 V2.9 software and a VC50 colour camera followed by image visualisation with HALO image analysis software.
RESULTS
Candidate genes important in SARS-CoV-2 infection are detectable at varying levels in human airway epithelial cells and lung tissue
We performed a targeted analysis of ACE2, TMPRSS2, ADAM17, CTSL, CD147, and GRP78 gene expression as candidates important for SARS-CoV-2 infection in human airway epithelial cells. Here and throughout gene expression analyses, CDH1 (E-cadherin) was used as a control for lung epithelial cell phenotype. We first examined these genes in a curated dataset of upper and lower airway epithelial cell gene expression from the nasal sinus to the 12th generation of airway in the lung (fig. 1a).

Microarray expression profiles of candidate SARS-CoV-2 receptor genes in upper and lower airways. a) Normalised log2 expression levels for ACE2, TMPRSS2, ADAM17, CTSL, CD147, and GRP78 genes compared across the upper airway (nasal, grey) and lower airways (trachea, green; large airway, blue; small airway, orange). CDH1 gene expression level is included as a positive control. b) Statistical values for comparisons for each gene at each airway generation. *=p<0.05; **=p<0.01; ***=p<0.001.
In the upper airways, all candidates were expressed with the highest levels observed for GRP78 and the lowest level observed for ACE2. Analysis along multiple generations of the lower airways (trachea, large (4–6th generation), and small airways (10–12th generation) revealed identical relative expression patterns with ACE2 being the least expressed and GRP78 being the highest expressed. ACE2 gene expression showed the greatest variability along the upper and lower airways, with greatest expression observed in the trachea samples and the lowest expression in the small airway (fig. 1b).
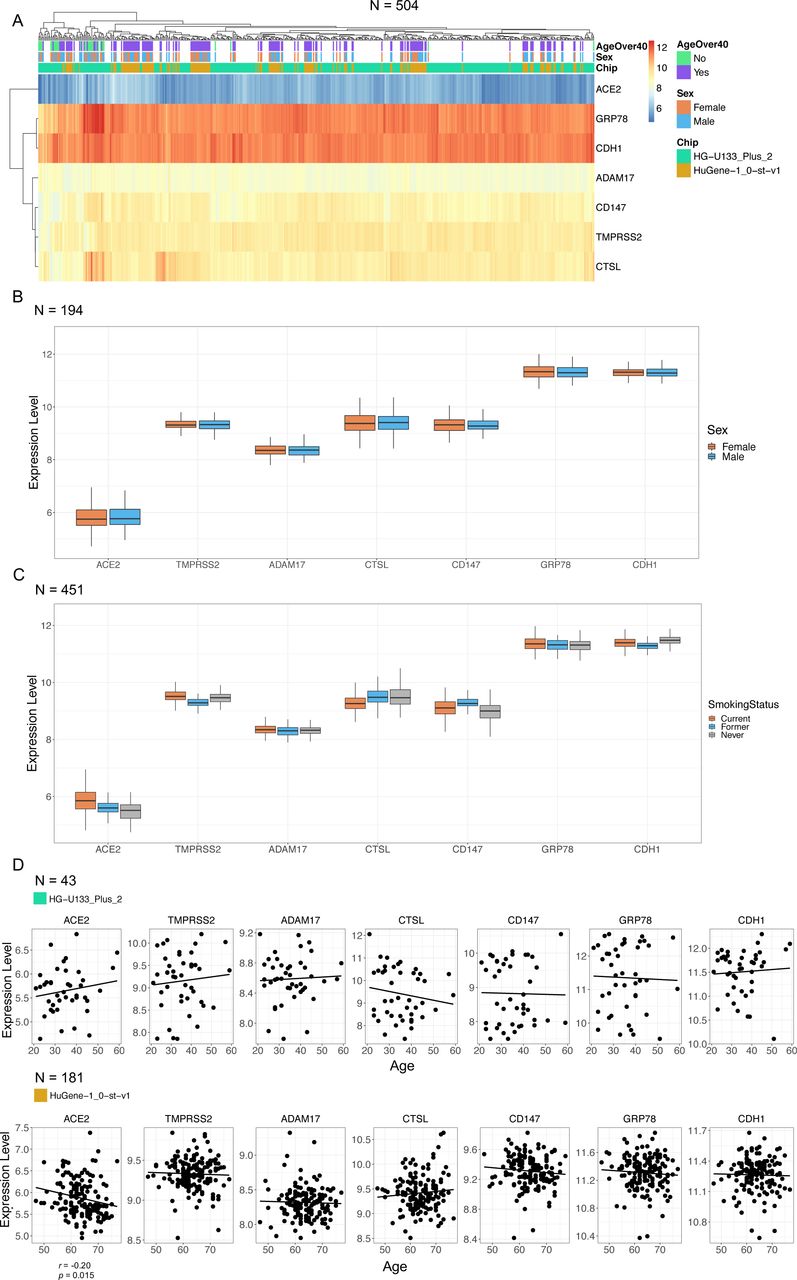
Following our observation of consistent expression along the upper and lower airways of candidate genes important in SARS-CoV, we determined if sex or age impacted gene expression levels in healthy individuals using a curated dataset of bronchial brushings from 504 healthy subjects (Supplement Table 1). The expression levels for the candidate genes in healthy subjects paralleled the patterns observed in the smaller survey of upper airways, trachea, large, and small airways (fig. 2). Median ACE2 gene expression was the lowest, while GRP78 gene expression was the highest (fig. 2a). No gene candidate demonstrated sex dependence for expression levels (fig. 2b). ACE2, TMPRSS2, CD147, and GRP78 were elevated in current smokers relative to never smokers (fig. 2c – Statistics in Supplement Table 2). CTSL was reduced in current smokers relative to never smokers. No microarray chip dependent effects were observed for relationships between sex or smoking status and gene expression. For quantitative analyses related to age and gene expression our curated database was divided into datasets that used either the HG-U133 Plus 2 or HuGene-1.0-st-v1 microarray due to differences in age distributions. In the HuGene-1.0-st-v1dataset (n=181) which included a greater proportion of older (>50) individuals, we observed reduced ACE2 gene expression with age (fig. 2d, p<0.05).

Microarray expression profiles of candidate SARS-CoV-2 receptor genes in lower airway epithelial cells analysed by age and sex. a) Clustered heatmap of log2 expression levels from 504 GEO samples, annotated by age, sex, and microarray chip platform. Expression values reflect signal intensities, indicating lowest detected expression of ACE2 and highest expression of GRP78 and CDH1. b) Per-gene boxplots of expression levels separated by sex. c) Per-gene boxplots of expression levels separated by smoking status. d) Plots of gene expression levels versus age, with linear regression lines of best fit. A weak negative correlation (r=−0.20, p=0.015) was detected for ACE2 in the second dataset. Correlations were performed separately between platforms because of differences in their age distributions.
Promoter activity data of each of the candidate genes important in SARS-CoV-2 binding and infection were extracted and analysed from the FANTOM5 dataset, which includes 1886 primary cells, cell lines, and tissue sample types (fig. 3). We selected all samples formats that included “lung”, “nasal”, “airway”, “olfactory” to identify lung-specific sample types. Gut, heart, and prostate tissue samples were analysed as controls. Consistent with our observed gene expression analysis along the upper and lower airways, normalised TPM values for each CAGE peak demonstrate that CD147 promoter activity was elevated relative to ACE2 promoter activity across airway epithelial cells and lung tissue samples. Cathepsin L promoter activity was the lowest of all candidate genes, which contrasted the modest expression observed at the gene level (fig. 2a). Both microarray gene expression analysis and promoter activity were consistent with results of candidate gene expression in a scRNAseq dataset of 10 healthy subjects (Supplementary Figure 1).

CAGE promoter activity for SARS-CoV-2-related genes from the FANTOM5 CAGE dataset. The FANTOM5 CAGE data consists of quantified promoter expression levels across the human genome for 1866 samples from primary cells, cell lines, and tissue samples. In this figure, the FANTOM5 CAGE promoter activity data for several SARS-CoV-2-related genes are shown for samples related to lung, gut, heart, and prostate tissues (n=120). Dot sizes are proportional to promoter activity, depicted as log10-transformed normalised transcripts per million (TPM). Notably, ACE2 is either not expressed or expressed at low levels (less than 1 TPM in all but one sample) in the airway, including measurements from healthy (white rows) and cancerous cells (grey rows).
Collectively, our gene expression analysis of the upper and lower airways of healthy males and females of diverse ages suggests that ACE2 gene expression is low relative to all other candidate SARS-CoV-2 receptor genes analysed in human airway epithelial cells. Furthermore, we observe no sex-dependent or age-dependent expression patterns of any candidates at the gene level, although smoking status impacted gene expression levels.
In vitro and in situ protein profiling reveals distinct expression patterns for candidates important in SARS-CoV-2 infection
Analysis of transcriptional data may not be indicative of in situ protein expression levels [49]. To extend our gene expression observations, we mined publicly available proteomic data from whole lung and primary human airway epithelial cell cultures and performed in vitro immunoblots on human airway epithelial cell lysates and in situ protein immunohistochemistry on human lung tissue using the same antibodies for each method.
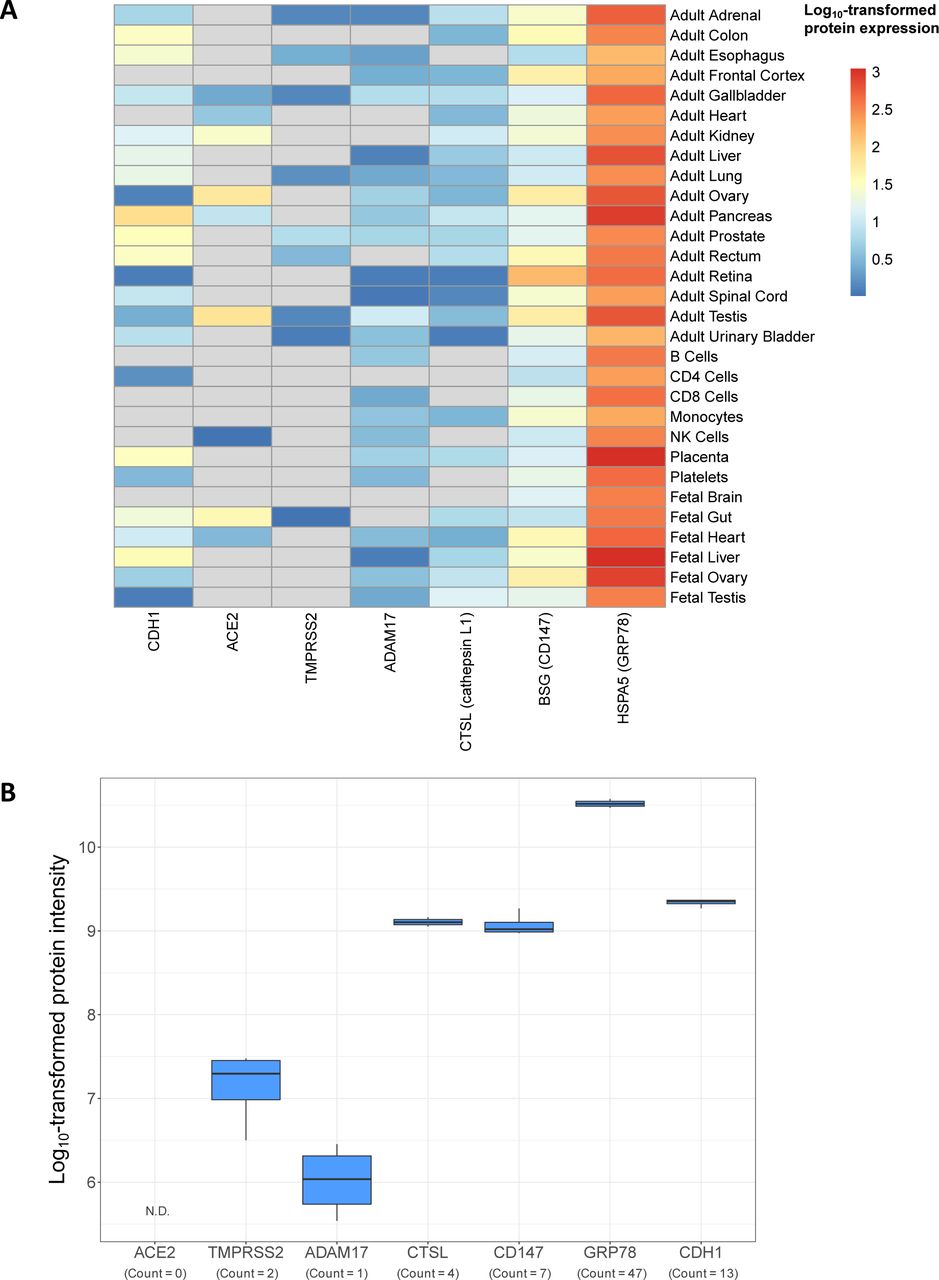
The Human Proteome Map is a publicly available resource that includes select adult and fetal tissues and circulating immune cell populations [31]. Using this resource, we examined protein expression of ACE2, TMPRSS2, ADAM17, CTSL, CD147, and GRP78. In human lung tissue homogenate, ACE2 was not detected, while being detected in heart, gut, and testes, known positive control tissues (fig. 4a). The rank order of the remaining molecules in human lung tissue homogenate was, GRP78>CD147>CTSL>ADAM17>TMPRSS2. Human lung tissue homogenate is a heterogeneous population of cells precluding the ability to associate protein expression to a given cell type. We therefore interrogated a publicly available proteomic dataset derived from primary human airway epithelial cells grown under air-liquid interface culture conditions [32], examining the same candidates. Again, ACE2 protein expression was not detectable (fig. 4b). CD147, GRP78, and CTSL were expressed with multiple peptide counts, while TMPRSS2 and ADAM17 were only marginally expressed with low peptide counts. Collectively, two proteomic datasets from distinct lung sample formats provide complementary and consistent expression profiles of candidate molecules important in SARS-CoV-2 infection.

Proteomic profiles of candidate SARS-CoV-2 receptor genes in human tissue and airway epithelial cells. a) Intensity values of protein expression from Kim et al. [31] for the genes ACE2, TMPRSS2, ADAM17, CTSL, CD147, and GRP78. CDH1 intensity is included as a positive control for expression in airway cells. Intensity values have been log10-transformed to facilitate comparison between candidates with different basal expression levels across tissue types. Grey cells in the heatmap correspond to an untransformed intensity of 0 and represent an undetectable signal. b) Intensity values log10-transformed for visualisation of ACE2, TMPRSS2, ADAM17, CTSL, CD147, and GRP78 proteins in human airway epithelial cells from healthy, non-smokers (n=4, males) grown under air-liquid interface culture conditions [32]. As in A, CDH1 intensity is included as a positive control. Counts indicating the number of detected peptides associated with each parent protein are provided. N.D. indicates the protein was not detected in this study.
To localise the in situ expression of the candidate molecules of interest at the protein level, we pursued immunohistochemical analysis paired with immunoblot validation of the specificity of the selected antibodies for recognition of proteins of predicted molecular weight. An anti-ACE2 antibody detected only a single band in Calu-3 cells at the predicted molecular weight (∼110 kDa) of ACE2 protein. The anti-ACE2 antibody required the use of a super-sensitive ECL solution (fig. 5a – Top panel - Lanes 1–3). No ACE2 protein was detected in primary airway epithelial cells or the HBEC-6KT cell line despite confirmation of protein loading (fig. 5a – Lanes 4–9, bottom panel confirms protein loading). Independent immunoblotting with a distinct anti-ACE2 primary antibody was performed, with a single band observed in HEK cells, but not in immortalised human bronchial epithelial cells or A549 cells (Supplement Figure 2).
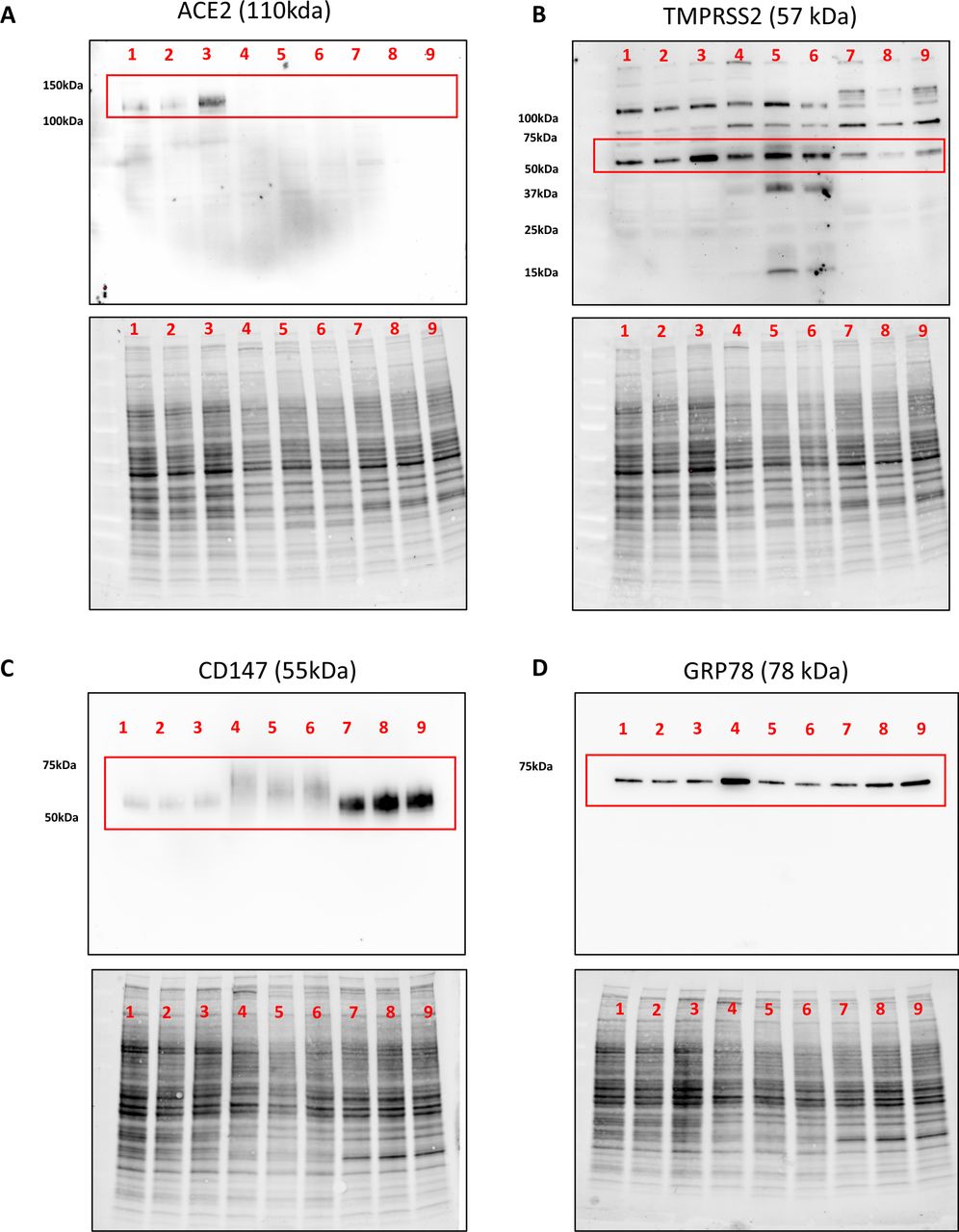
Immunoblot analysis of ACE2, TMPRSS2, CD147, and GRP78 protein expression in human airway epithelial cell protein lysates. a) ACE2 with single band for predicted molecular weight of 110 kDa (red box). b) TMPRSS2 with multiple bands including a dominant band at predicted molecular weight of 57 kDa (red box). c) CD147 with a single broad band around predicted molecular weight of 55 kDa (red box). d) GRP78 with a single band at predicted molecular weight of 78 kDa (red box). Lanes 1–3: Calu-3 cells. Lanes 4–6: Primary human airway epithelial cells. Lanes 7–9: HBEC-6KT. All cells grown under submerged monolayer conditions, with n=3 independent passages (Calu-3 or HBEC-6KT) or donor samples (Primary human airway epithelial cells – non-smoker, healthy subjects). For each independent blot of each protein, all of the same samples were run. Total protein loading control provided to demonstrate protein loaded for each sample.
An anti-TMPRSS2 antibody detected multiple bands in all airway epithelial cell samples with a dominant band at the predicted molecular weight of ∼55 kDa (fig. 5b – Top panel). These patterns were conserved across all cell types that were analysed.
An anti-CD147 antibody detected a single band in all airway epithelial cell samples with a dominant band at the predicted molecular weight of ∼55 kDa (fig. 5c – Top panel). The immunoblot bands were consistent with the heavy glycosylation of CD147 [50].
An anti-GRP78 antibody (BD - 610 979) detected a single band in all airway epithelial cell samples with a dominant band at the predicted molecular weight of ∼78 kDa (fig. 5d – Top panel).
The immunoblots using anti-ACE2, CD147, and GRP78 demonstrated a single band of predicted molecular weight, suggesting that observed immunohistochemical staining should be specific to the protein of interest based on the target epitope, as both methods detect denatured proteins [51]. The same anti-ACE2 and CD147 antibodies were validated for immunohistochemistry. The anti-TMPRSS2 was used for immunohistochemistry, although the multiple bands observed by immunoblot caution the specificity of any observed in situ staining. Attempts to optimise anti-GRP78 antibody application for immunohistochemistry were unsuccessful, requiring additional antibody interrogation with HPA038845 (Atlas Antibodies) that was suitable for both immunoblotting and immunohistochemistry (Supplement Figure 3).
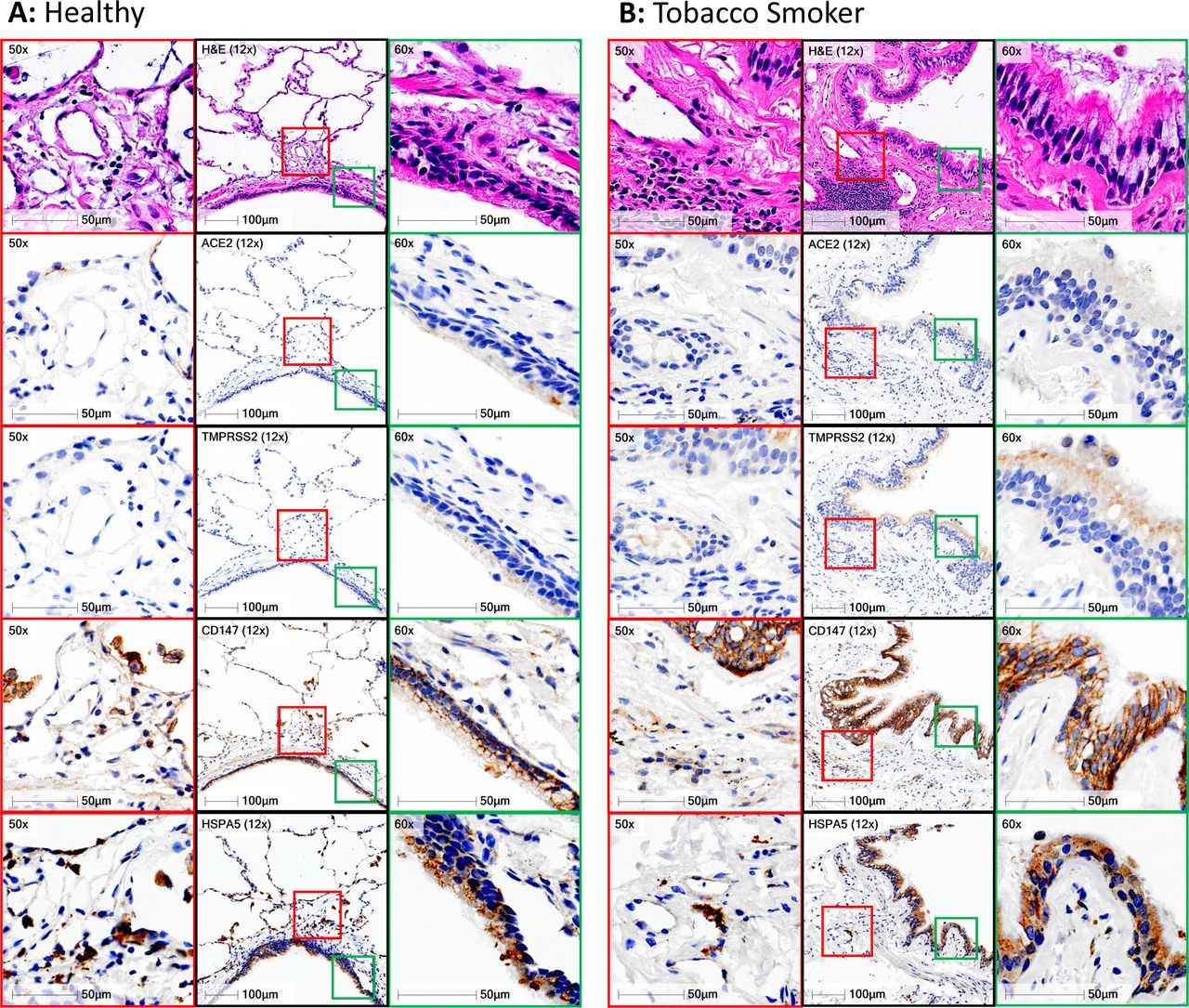
ACE2 immunohistochemistry revealed only select staining in rare cells in the airways and the alveoli of all 98 human lung samples analysed that included healthy subjects and those with chronic lung diseases (fig. 6). A single healthy human sample contained one positive airway epithelial cell with additional positive staining in the peripheral lung in cells with type II alveolar epithelial cell morphology (fig. 6a – second row). A representative image of a sample from a smoker with chronic obstructive pulmonary disease (fig. 6b – second row) shows no ACE2 protein staining in the airway epithelium and a rare positive cell in sub-basement membrane tissue. Quantification of positive pixel count for ACE2 staining normalised to total tissue pixel count revealed no differences between healthy non-smokers and tobacco smokers (Supplement Figure 4). Lung microvasculature and human heart tissue stained had positive staining (Supplementary Figures 5–6) consistent with previously described reports for ACE2 protein staining patterns [52, 53].

Immunohistochemical localisation of ACE2, TMPRSS2, CD147 and GRP78 protein in human lung tissue. a) Representative panel of a healthy non-smoker with no underlying chronic airway disease. b) Representative panel of a smoker with chronic obstructive pulmonary disease. For both panels, black squares represent low magnification (12X) of a conducting airway with airway epithelium. Green squares correspond to high magnification regions (50X) of conducting airway epithelium that are defined in the low magnification image. Red squares correspond to high magnification regions (50X) of lung tissue away from airway lumen that are defined in the low magnification image. Row 1 - hematoxylin & eosin, Row 2 – ACE2, Row 3 – TMPRSS2, Row 4 – CD147, Row 5 – GRP78/HSPA5. Positive immunohistochemical staining is rust/brown. Total number of independent samples analyses was 49–98.
TMPRSS2 immunohistochemistry revealed diffuse staining in the airway epithelium and in immune cells in the lung periphery, with greater staining in smokers with chronic obstructive pulmonary disease (COPD) (fig. 6a and b – third row). These observations were consistent in the majority of the 98 human samples examined.
CD147 immunohistochemistry revealed strong membrane restricted staining in the airway epithelium and diffuse staining in immune cells in the lung periphery (fig. 6a and b – fourth row). CD147 displayed greater staining in smokers with COPD. These observations were consistent in the majority of the 98 human samples examined.
GRP78 immunohistochemistry revealed diffuse staining in airway and alveolar epithelium and in immune cells in the lung periphery (fig. 6a and b – fifth row). No qualitative differences in GRP78 staining were observed between healthy subjects and smokers with COPD.
Collectively, our in vitro and in situ protein profiling is consistent with our gene expression analysis with CD147 and GRP78 protein expression dominant over TMPRSS2 and ACE2. Additional examples of staining in lung tissue are provided in Supplement Figure 7. ACE2 protein expression is rare in human lung tissue and found in select cells in both healthy individuals and those with chronic lung diseases. TMPRSS2 and CD147 protein expression are potentiated in individuals with a history of tobacco smoking and a diagnosis of COPD.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
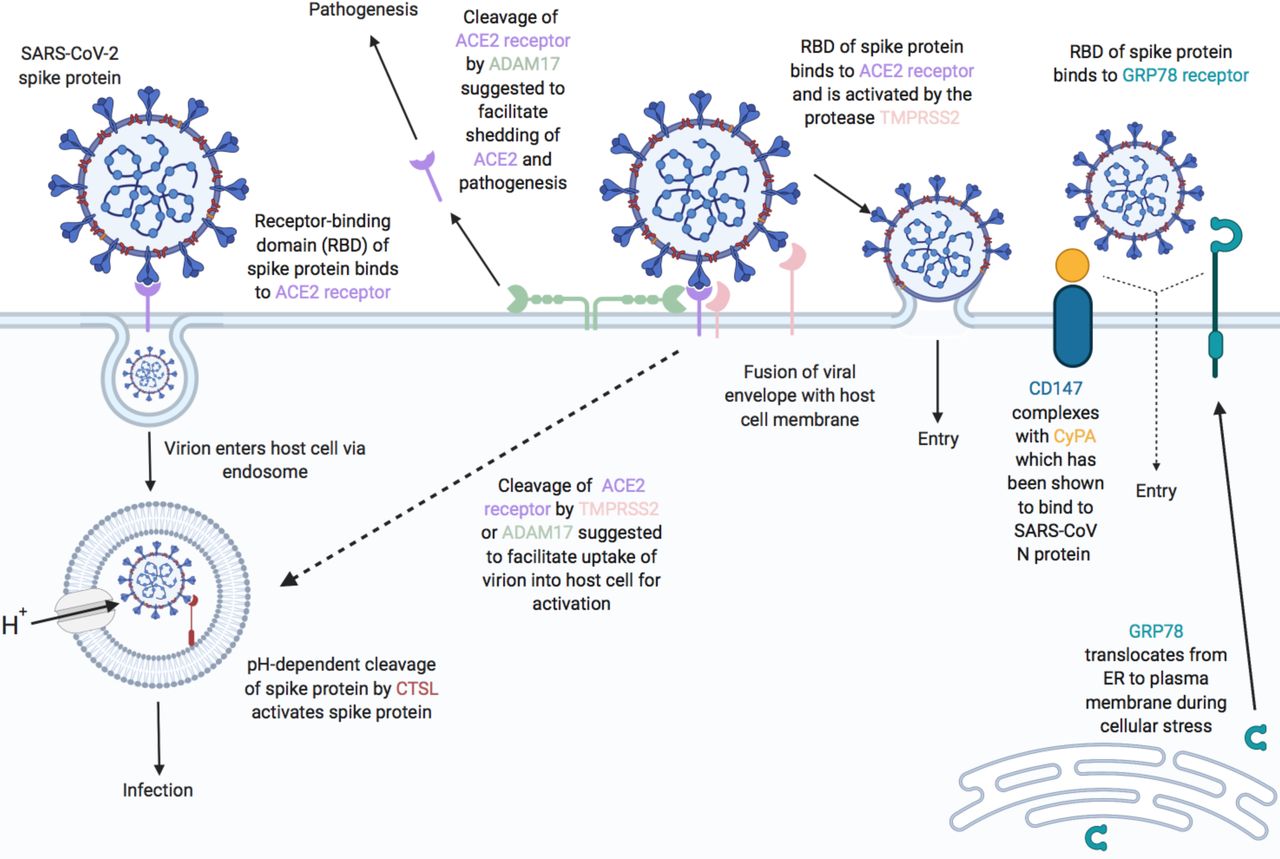
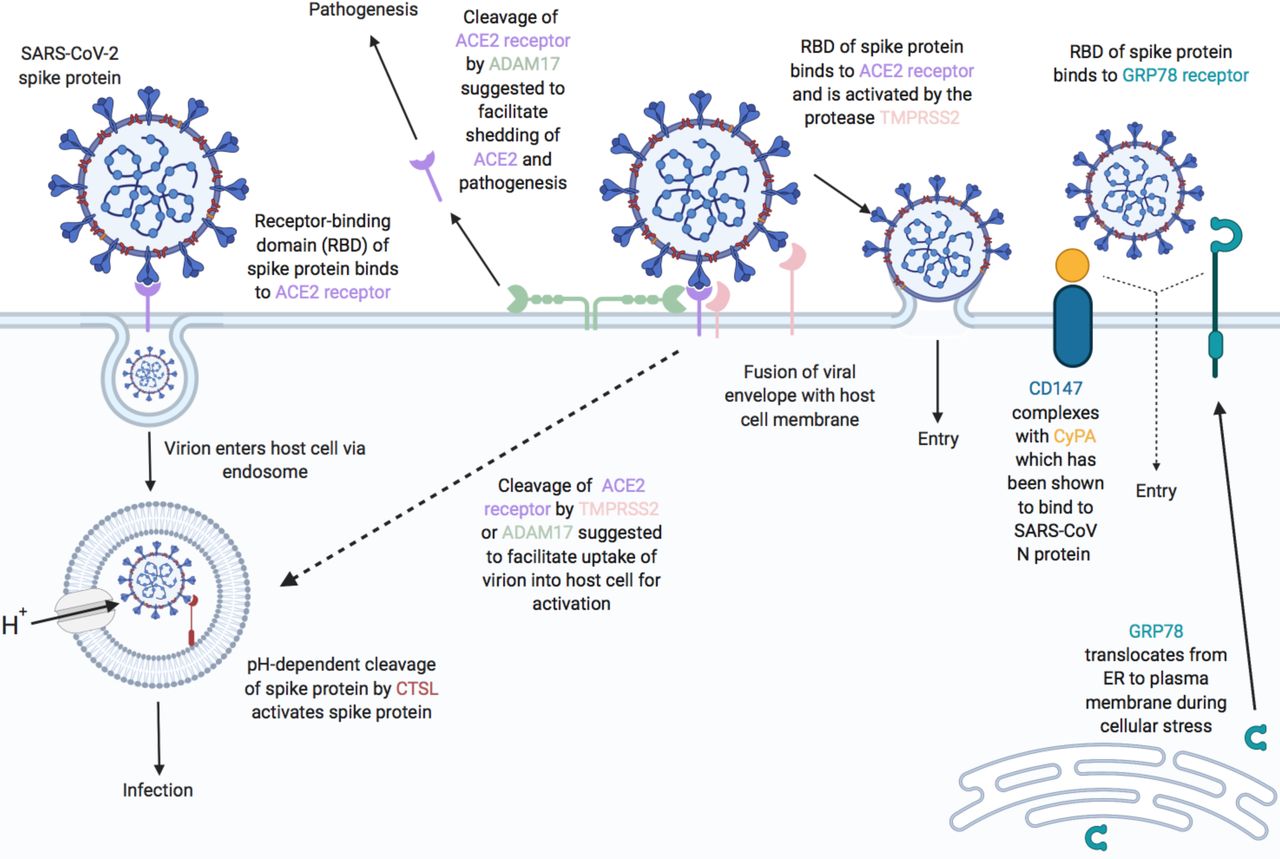
Proposed functions of host airway epithelial cell molecules for interaction with SARS-CoV-2. Proteins associated (or suggested to be associated) with host cell entry of SARS-CoV-2 and the activation of the SARS-CoV-2 spike protein (SARS-S) are displayed. ACE2 is suggested as the primary SARS-S receptor for viral entry (interaction of ACE2 receptor-binding domain (RBD) with SARS-S leading to endosomal viral uptake) followed by activation of SARS-S via pH-dependent CTSL-mediated cleavage. Secondary methods of viral entry and SARS-S activation are likely to involve proteases (e.g. TMPRSS2 and ADAM17) and/or secondary receptors (CD147 and GRP78). Dashed lines indicate mechanisms that have not been fully validated. Figure adapted from [20] with updates and additional information on candidate host molecules. Created with BioRender.com.
DISCUSSION
The global COVID-19 pandemic that emerged in late 2019 is caused by SARS-CoV-2. The possible host receptor(s) for SARS-CoV-2 have not been exhaustively surveyed in human lung tissue at the gene and protein level. Understanding the expression levels and localisation of candidate SARS-CoV-2 receptors in host lung tissue may provide insights into therapeutic interventions that might reduce disease spread, viral replication, or disease pathology. To address this knowledge gap, we performed gene expression, proteomic profiling at the tissue and cell level, and in situ protein profiling of candidate receptors in human airway epithelial cells and lung tissue (summarised in fig. 7). Collectively our data demonstrate rare ACE2 protein expression in human airway epithelial cells in vitro and in situ. Our protein expression data are consistent with low ACE2 promoter activity in a panel of lung epithelial cell samples and low ACE2 gene expression in bronchial epithelial cells (microarray) and lung cells (scRNAseq). We present confirmatory evidence for the presence of TMPRSS2, CD147, and GRP78 protein in vitro in airway epithelial cells and confirm broad in situ protein expression of CD147 and GRP78 in the respiratory mucosa. Our data suggest that for ACE2 to be an integral receptor for SARS-CoV-2, mechanisms are likely to exist that dynamically regulate expression in human lung, perhaps in periods of SARS-CoV-2 infection [54]. It is also possible that alternate receptors for SARS-CoV-2 are important in initial host cell infection.
Using a curated microarray gene expression dataset generated from bronchial brushings of 504 healthy subjects that considers the limitations of merging multiple datasets from distinct sources, we observed that sex did not correlate with gene expression of any candidate host molecule involved in SARS-CoV-2 infection and that ACE2 and TMPRSS2 were the lowest expressed genes of interest examined. In one dataset, ACE2 gene expression modestly decreased with age, although protein level confirmation was not possible. The low level of ACE2 and TMPRSS2 gene expression in bulk bronchial epithelial cell gene expression samples suggests low levels of cells expressing both of these genes within this lung tissue. We confirm that tobacco smoking is associated with elevated ACE2 gene expression levels in bronchial epithelial cell samples [55], although we were unable to confirm this with immunohistochemistry analysis of protein on human lung samples.
Advances in transcriptomics have enabled scRNAseq that has identified unique and rare cell types in human lung that may have importance in health and disease [56, 57]. scRNAseq provides an opportunity to look at transcriptional profiles in subsets of cell populations, which may isolate a cell signal from a bulk sample. We therefore utilised scRNAseq data from healthy human lung samples as a parallel approach. The resolution of scRNAseq for subpopulations of epithelial cells revealed low or absent expression of ACE2 gene in all populations examined, whereas CD147 and GRP78 were present in all populations. Our results are consistent with current publicly available data that discuss the presence of a rare ACE2/TMPRSS2 positive cells [54]. Using lung samples from eight individuals (four HIV and active tuberculosis double +ve, two HIV +ve and tuberculosis -ve, and two double -ve controls), Ziegler et al. have reported in humans that only 0.8% of type II alveolar epithelial cells expressed both ACE2 and TMPRSS2 genes [54]. Further analysis of ciliated cells found that 5.3% of these cells expressed both ACE2 and TMPRSS2 genes. In vitro models with SARS-CoV, are consistent with this finding as ciliated cells are preferentially targeted by this coronavirus [58]. Most intriguing is that ACE2 and TMPRSS2 gene expressing cells were only identified in the HIV and tuberculosis double +ve samples. These observations were replicated in the upper airways, with only a rare population of secretory epithelial cells (0.3% of this population) co-expressing ACE2 and TMPRSS2. The reported scRNAseq results are consistent with a focused analysis looking at only ACE2 gene expression in a variety of lung cell types [59]. Importantly, these elegant transcriptomic analyses confirm our observations in bulk tissue microarray datasets.
Consortium based publicly available datasets represent another parallel approach to confirm our data. We have used the FANTOM5 dataset containing CAGE promoter activation data for 1866 primary cells, cell lines, and tissue samples from humans [39] to examine the level of promoter activity for each candidate SARS-CoV-2-receptor genes. The FANTOM5 CAGE data provides an additional and complementary approach to quantifying gene expression since a given gene's shared promoter can yield multiple transcripts at different expression levels, as well as being partially independent of any given transcript's half-life in the cell. In general, the promoter activity of ACE2 in airway-related tissues is low or absent; only a single sample originating from an adult lung yields a normalised CAGE promoter expression level above one transcript per million, while expression was observed in gut cells, consistent with known patterns of ACE2 expression [60]. Consistent with the microarray data, CD147 promoter activity is elevated relative to ACE2 across airway-related cells and tissues, although the relatively low CTSL (cathepsin L1) promoter activity is incongruent with modest levels of gene expression.
The expression of genes does not always correlate with protein expression [49]. With this in mind we performed combination proteomic analyses with immunoblot analyses. For our immunoblots, we used the human Calu-3 adenocarcinoma cell line as this cell is susceptible and permissive to SARS-CoV-2 infection and expresses ACE2, an observation we confirm [24, 29]. We also used primary human airway epithelial cells and the bronchial epithelial cell line (HBEC-6KT). We performed immunoblots for ACE2 and TMPRSS2 as these have been highlighted as interacting with SARS-CoV-2, while we probed CD147 as recent pre-clinical and clinical studies have provided proof of concept as a candidate SARS-CoV-2 receptor [26, 27]. Lastly, GRP78 was dominantly expressed throughout transcriptomic studies and was selected as a positive control as previous expression has been confirmed in human airway epithelial cells [61]. Cathepsin L was excluded from the present analysis due to low promoter activity (fig. 3), while ADAM17 was excluded as the proposed function in coronavirus infections is via ACE2 [5, 24], which was included in analysis. Immunoblot analysis with all antibodies revealing dominant bands of predicted molecular weight, with the anti-TMPRSS2 polyclonal antibody revealing additional minor bands in all cell samples examined. The identity of these other bands remains unclear and suggest downstream immunohistochemical analysis may be confounded by the specificity of this antibody. In contrast, antibodies for ACE2, CD147, and GRP78 were specific and could be used for immunohistochemistry without concerns of specificity. Interestingly, ACE2 protein could only be detected with a supersensitive ECL solution and only in Calu-3 cells, suggesting absent protein in primary human airway epithelial cell and the HBEC-6KT cell lines. Our data are consistent with previous immunoblots of primary human airway epithelial cells grown under submerged monolayer conditions using the same primary antibody, where ACE2 protein was absent, and only expressed under air-liquid interface culture conditions [62]. The observation that CD147 and GRP78 are also expressed in Calu-3 cells encourages further interrogation into these host proteins, as they may contribute to function of ACE2 and TMPRSS2 in SARS-CoV-2 binding and fusion in this cell type. Collectively, the profiling of antibodies by immunoblot of airway epithelial cells revealed distinct band patterns demonstrative of antibody specificity for ACE2, CD147, and GRP78, and to a lesser extent for TMPRSS2.
Immunohistochemical analysis has been performed for localisation of ACE2 and TMPRSS2 in human lung [28, 30]. The observation of positive staining in human lung tissue for these proteins was not accompanied by companion immunoblot or complementary approaches to define the specificity of the antibody used [51]. In the absence of determination of antibody specificity, the historical data presented should be interpreted with caution. To address the issue of antibody specificity for immunohistochemical staining, we used the same antibodies we validated by immunoblot and confirmed findings using proteomics as an orthogonal, antibody independent, approach. We again focused on ACE2 and TMPRSS2 as these are candidate proteins important for SARS-CoV-2 infection of host cells. Our immunohistochemical staining patterns of ACE2 were consistent with transcriptional profiling and immunoblots with only 1 of 98 human samples demonstrating rare staining in the airway and alveolar epithelium. Positive ACE2 staining in heart tissues and areas of lung microvasculature suggest our staining protocol was successful. These results directly contrast those reported with antibodies that lacked validation for specificity [28, 30]. TMPRSS2 was expressed more frequently across all samples examined with variability in the airway epithelium, associated with history of smoking and/or chronic obstructive pulmonary disease status. In contrast, CD147 expression was observed in airway epithelium of all samples. Similar to TMPRSS2, elevated CD147 expression was associated with history of smoking and/or chronic obstructive pulmonary disease status, consistent with previous reports [50]. Our original GRP78 antibody selected for immunoblotting was not validated for immunohistochemistry. We therefore performed confirmatory GRP78 immunoblotting and immunohistochemistry with an additional antibody (HPA038845) and provide demonstration of expression in human airway epithelial cells in vitro and in situ. Importantly, it is well established that GRP78 resides in the endoplasmic reticulum (ER) under normal physiological conditions where it acts as an ER-resident molecular chaperone to facilitate correct protein folding. However, under conditions of ER stress, including viral infection, a portion of ER-resident GRP78 relocates to the cell surface where it may act as a viral co-receptor [63]. Our research group has reported the presence of cell surface GRP78 in atherosclerotic plaques [64], prostate cancer [65] and kidney [66]. However, there are currently no commercially available anti-GRP78 antibodies that bind specifically to cell surface GRP78 and dual immunofluorescence is used to show colocalization of cell surface GRP78 with an established surface receptor [64]. This drawback precludes our ability to perform accurate cell surface GRP78 immunohistochemistry on lung tissue to interrogate this concept further in the context of SARS-CoV-2 receptors. Utilisation of prostate cancer patient-derived GRP78 auto-antibodies that are specific for cell-surface GRP78 [64] may be suitable on lung tissue for assessment of cell susceptibility to SARS-CoV-2 infection by GRP78.
Our study has several limitations that have not already been addressed above. Our observation of differences in gene expression between upper and lower airways and along the airway tree were not corroborated at the protein level. It remains possible that entirely different protein expression profiles for the candidate molecules examined exist in the upper airway, presenting a different environment for SARS-CoV-2 interaction with the respiratory mucosa. Nasal pharyngeal swabs are capable of detecting SARS-CoV-2 virus [67] and this anatomical region likely is important for subsequent infection in the lower airways [8, 68]. Related to this potential temporality of effect, it is possible that SARS-CoV-2 induces the expression of receptors on host cells following infection [54]. Our study is also limited by examining candidate molecules important in SARS-CoV-2 infection under basal conditions, in the absence of viral or environmental stimuli which may regulate gene transcription and protein translation.
SARS-CoV-2 infection and transmission has caused the global COVID-19 pandemic. An understanding of the receptors used by SARS-CoV-2 for host cell infection and the parallel characterisation in human samples is required to inform development of intervention strategies aimed at mitigating COVID-19. Our data demonstrate rare ACE2 protein expression in human airway epithelial cells in vitro and in situ, consistent with low ACE2 promoter activity and ACE2 gene expression in bronchial epithelial cells. We present confirmatory evidence for the presence of TMPRSS2, CD147, and GRP78 protein in vitro in airway epithelial cells and confirm broad in situ protein expression of CD147 and GRP78 in the respiratory mucosa. Due to the overwhelming evidence that SARS virus interacts with ACE2, there are likely to be alternate mechanisms regulating ACE2 in the respiratory mucosa in the context of SARS-CoV-2 infection, and/or, perhaps other co-receptors, beyond what is expressed under basal conditions at the protein level.
Acknowledgements
We would like to acknowledge Mary Jo Smith from the McMaster Immunology Research Centre Core Histology facility for her timely and professional expertise with antibody staining for immunohistochemistry. We would like to thank Drs. Sam Wadsworth and John McDonough for their intellectual discussions around CD147 and airway epithelial cell biology. We would like to thank Dr. Charles Plessy for his suggestion to analyse the FANTOM5 data. We would like to thank all of the personal and professional support from the author's respective families (CLB, JMH, and HLH) and research institutes, and most importantly, the frontline healthcare workers during the COVID-19 pandemic.
Footnotes
This article has supplementary material available from erj.ersjournals.com
Support statement: This work was supported by start-up funds from Hirota JA and a CIHR grant from Mossman K. Hirota JA is supported by the Canada Research Chairs program and an Ontario Early Researcher Award. Doxey AC is supported by NSERC and an Ontario Early Researcher Award. Canada Research Chairs; DOI: http://dx.doi.org/10.13039/501100001804; Canadian Institutes of Health Research; DOI: http://dx.doi.org/10.13039/501100000024.
Conflict of interest: Dr. Aguiar has nothing to disclose.
Conflict of interest: Dr. Tremblay has nothing to disclose.
Conflict of interest: Dr. Mansfield has nothing to disclose.
Conflict of interest: Dr. Woody has nothing to disclose.
Conflict of interest: Dr. Lobb has nothing to disclose.
Conflict of interest: Dr. Banerjee has nothing to disclose.
Conflict of interest: Dr. Chandiramohan has nothing to disclose.
Conflict of interest: Dr. Tiessen has nothing to disclose.
Conflict of interest: Dr. Cao has nothing to disclose.
Conflict of interest: Dr. Dvorkin-Gheva has nothing to disclose.
Conflict of interest: Dr. Revill has nothing to disclose.
Conflict of interest: Dr. Miller has nothing to disclose.
Conflict of interest: Dr. Carlsten has nothing to disclose.
Conflict of interest: Dr. Organ has nothing to disclose.
Conflict of interest: Dr. Joseph has nothing to disclose.
Conflict of interest: Dr. John has nothing to disclose.
Conflict of interest: Dr. Hanson has nothing to disclose.
Conflict of interest: Dr. Austin has a patent US7524826B2 issued to MCMASTER UNIVERSITY AND HAMILTON HEALTH SCIENCES Corp McMaster University.
Conflict of interest: Dr. McManus has nothing to disclose.
Conflict of interest: Dr. Jenkins reports grants from Astra Zeneca, grants from Biogen, personal fees from Boehringer Ingelheim, personal fees from Daewoong, personal fees from Galapagos, grants from Galecto, grants from GlaxoSmithKline, personal fees from Heptares, non-financial support from NuMedii, grants and personal fees from Pliant, personal fees from Promedior, non-financial support from Redx, personal fees from Roche, other from Action for Pulmonary Fibrosis, outside the submitted work.
Conflict of interest: Dr. Mossman has nothing to disclose.
Conflict of interest: Dr. Ask has nothing to disclose.
Conflict of interest: Dr. Doxey has nothing to disclose.
Conflict of interest: Dr. Hirota has nothing to disclose.
- Received April 10, 2020.
- Accepted July 1, 2020.
- Copyright ©ERS 2020
This article is open access and distributed under the terms of the Creative Commons Attribution Non-Commercial Licence 4.0.
References










